
Er DNA-replikation dårligt designet?
Alle celler bruger DNA som genetisk materiale, og ved celledeling kopieres DNA’et. Denne proces, kaldet DNA-replikation, involverer en række specialiserede proteiner og er, fra et design-perspektiv, et godt bud på et intelligent designet biologisk system. Dog er der et aspekt ved DNA-replikationen, den såkaldte følgestrengssyntese (på engelsk, lagging strand syntesis), som er blevet kritiseret som værende udtryk for dårligt design.
I denne artikel vil jeg kigge nærmere på denne kritik og vise, hvorfor der tværtimod er grund til at se dette aspekt ved DNA-replikationen som udtryk for godt design. Kritikken er illustrativ, da den viser designtesens forklaringskraft.
Replikationsmekanismen
Inden en celle kan deles og blive til to, må dens DNA kopieres. DNA’et er en dobbeltstrenget spiral, og når replikationsprocessen starter, går specialiserede proteiner i gang med at adskille disse to strenge.
DNA’ets byggesten er nukleotider, og hver streng består af en lang kæde af disse nukleotider, hæftet sammen med fosfatbindinger. Nukleotiderne findes i fire forskellige varianter (forkortet A, T, C og G), og det er rækkefølgen af disse varianter, der koder for den information, cellen bruger til at producere proteiner.
De to DNA-strenge er bundet sammen af hydrogenbindinger, der dannes mellem hvert nukleotide-par. De fire nukleotide-varianter binder sig til hinanden parvis, således at A binder sig til T og C binder sig til G (figur 1).
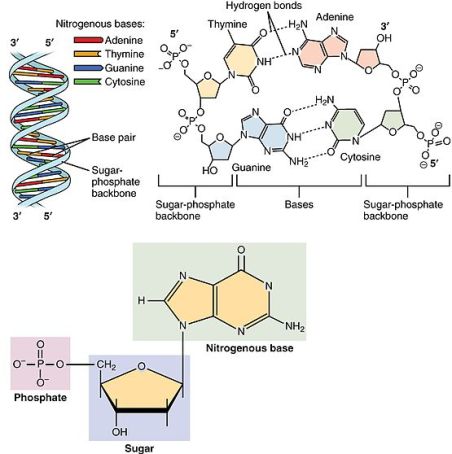
Denne parvise binding betyder, at hver streng er et spejlbillede, eller negativ af den anden streng. Og at hver streng således indeholder den nødvendige information til at danne dens modstykke.
Efterhånden som de to DNA-strenge adskilles, går et særligt protein kaldet DNA-polymerase i gang med at kopiere dem. Et polymerase-protein bevæger sig langs med hver streng, som kopieres, hvorved der dannes to dobbeltstrengede kopier af DNA’et.
Den kluntede følgestrengssyntese?
De to strenge kopieres imidlertid ikke på helt samme måde.
Nukleotider har polaritet, forstået på den måde, at der er forskel på den ene og den anden ende. Nukleotidet er opbygget af fem karbonatomer, og disse er nummererede fra 1′ til 5′ (figur 2).
Fosfatbindingen dannes mellem 3′-atomet på et nukleotid og 5′-atomet på et andet. Man kan forestille sig nukleotiderne som Lego-klodser stablet oven på hinanden, hvor oversiden på en Lego-klods er sat sammen med undersiden på en anden.
Dette betyder også, at hver DNA-streng har polaritet, og man skelner mellem 3′-enden og 5′-enden.
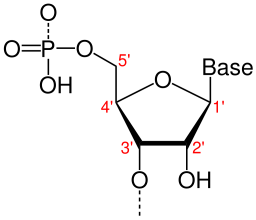
De to DNA-strenge er antiparallelle, det vil sige, at de vender hver sin vej; man kan forestille sig to stabler af Lego-klodser, hvor den ene stabel vender knopperne op og den anden stabel vender knopperne ned.
DNA-polymerasen bevæger sig imidlertid kun i én retning: Fra 5′-enden mod 3′-enden. Det betyder, at på den ene DNA-streng, kaldet lederstrengen, vil DNA-polymerasen bevæge sig i samme retning, som DNA’et adskilles. Men på den anden streng, kaldet følgestrengen, vil DNA-polymerasen bevæge sig i den modsatte retning. Dermed replikeres lederstrengen jævnt, mens følgestrengen replikeres i afbrudte etaper, da DNA-polymerasen hele tiden er nødt til at hoppe tilbage (figur 3).
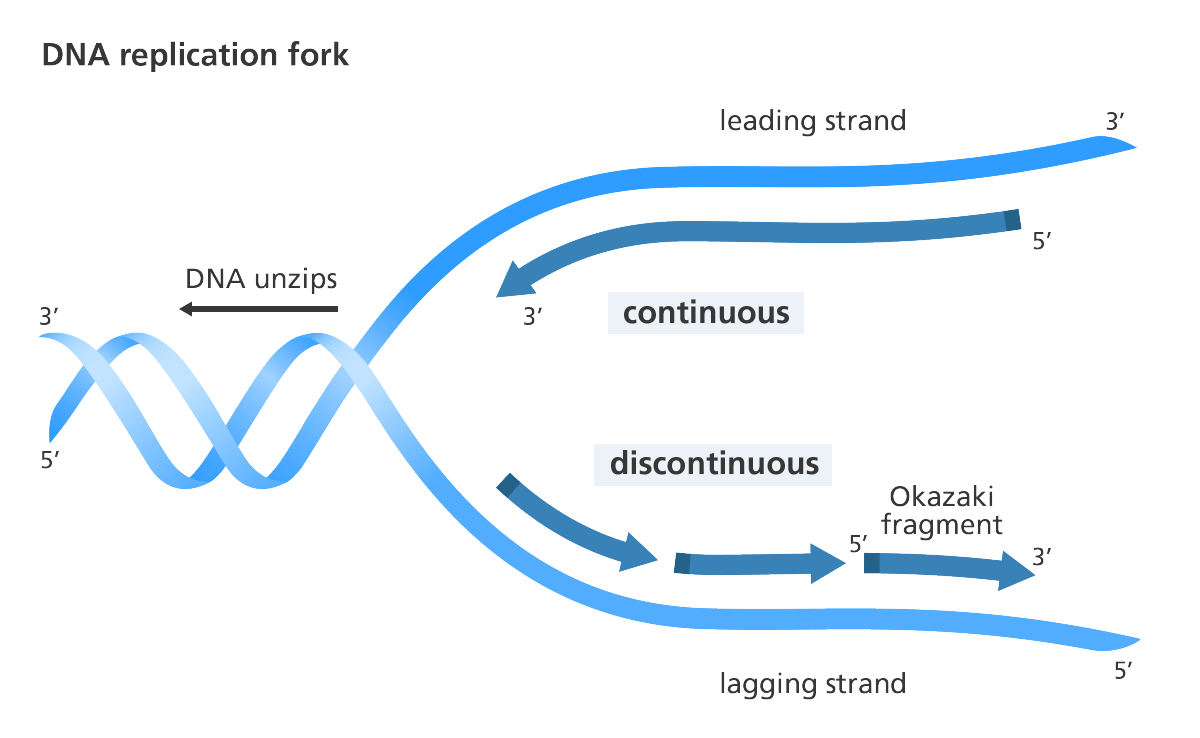
Dette aspekt af DNA-replikationen er blevet betegnet som værende udtryk for dårligt design:
“[Følgestrengssyntesen er] en meget kluntet løsning, eller i hvert fald en løsning, der er langt mere besværlig, end en, et menneske ville forsøge at designe. … Et menneske, der forsøgte at designe en løsning, ville formentlig foreslå, at man simpelthen lavede en anden type DNA-polymerase, der kunne læse DNA i den modsatte retning. Der er trods alt intet magisk ved retningen. I princippet burde det kunne gøres ved at foretage ændringer af DNA-polymerasen. Naturens løsning på problemet er dog en komedieserie værdig.” (Rensberger, 1996, s. 128-9)
Hvis DNA-replikationen er dårligt designet, ville dette være svært at forene med en designtese. Cellens maskineri er mere avanceret end noget, vi mennesker kan designe, og en eventuel designer må derfor formodes at have været mindst lige så intelligent som os. Hvis vi kan identificere en fejl ved designet, må en designer også have været i stand til at identificere denne fejl og rette den.
Designtesen lader os derfor fremsætte en forudsigelse: Enten er den pågældende struktur alligevel ikke designet, eller også er den alligevel ikke udtryk for dårligt design. Som jeg vil vise i næste sektion, er det sandsynligvis det sidste, der gør sig gældende.
Den elegante følgestrengssyntese
Syntesen af den replikerede DNA-streng er en energikrævende proces. Når cellen producerer nukleotider, producerer den dem i en “aktiveret” form, med tre fosfatmolekyler i 5′-enden. Når DNA-polymerasen tilføjer et nukleotid, fraspaltes to af disse fosfatmolekyler, hvilket frigiver energi og danner den sukker-fosfat-kæde, der udgør rygraden i DNA-strengen (figur 4).
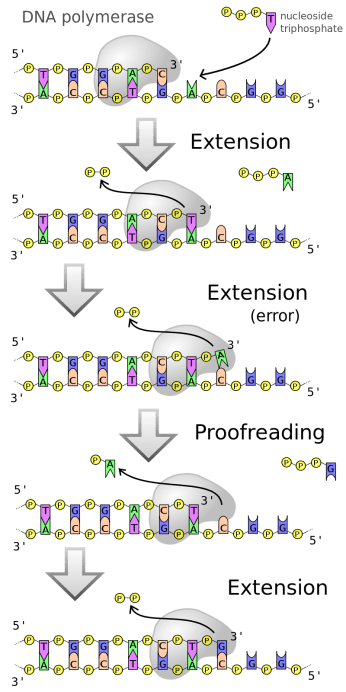
Når DNA-polymerasen bevæger sig i 5′-til-3′-retningen (som den gør i alle eksisterende celler), tilføjes nye nukleotider således med den fosfatbærende 5′-ende vendt mod den fosfatløse 3′-ende på det sidst tilføjede nukleotid (figur 4, øverst).
Hvis vi forestiller os, at vi forsøgte at “fikse” følgestrengssyntesen ved at designe en DNA-polymerase, der bevægede sig i 3′-til-5′-retningen, ville det derimod være det sidst tilføjede nukleotid, hvis fosfatbærende 5′-ende indeholdt energien til tilføjelsen af det næste nukleotid. Dette ville betyde, at hvis DNA-polymerasen kom til at tilføje et forkert nukleotid, som derefter skulle fjernes, ville dette også fjerne fosfatmolekylerne, og DNA-replikationen ville gå i stå.
Som en lærebog i cellebiologi formulerer det:
“Korrekturlæsningen kan forklare, hvorfor DNA-syntesen kun sker i 5’→3′-retningen. Ved nedlæggelse i 5’→3′-retningen kan fejlindbygning af sidst tilkommende nukleotid rettes ved simpelthen at fraspalte det igen. Den efterfølgende indbygning af et korrekt nukleotid giver ingen problemer, idet dette som nævnt selv bærer den triphosphatgruppen [sic], der er nødvendig for dannelsen af den covalente binding. Forestiller man sig imidlertid en syntese i 3’→5′-retning, er 5′-trifosfaten i det allerede nedlagte nukleosidstriphosphat, der bindes til næste nukleotids 3′-OH-gruppe. Fjernelse af et forkert nukleotid vil medfører [sic], at strengen terminerer 3′-OH-gruppen i stedet for den nødvendige triphosphatgruppe, og videre DNA-syntese er dermed forhindret.” (Prentø & Jensen, 2000, s. 234-5)
Følgestrengssyntesen er således ikke en “kluntet” løsning, som en intelligent designer ville have undgået. Det er derimod en ganske elegant løsning, som sikrer, at DNA-replikationen kan fortsætte, selv hvis der sker fejl.
Designtesens forklaringskraft
Kritikere af intelligent design hævder ofte, at konceptet er et tomt “designeren gjorde det”-udsagn, der kan forklare enhver tænkelig observation og derfor intet forklarer.
Jeg er uenig i denne kritik. Intelligent design giver os en berettiget forventning om, at designede strukturer lever op til empirisk identificérbare kriterier for “godt design”. Intelligent design giver os således bestemte forventninger til, hvad vi kan observere i naturen.
Der er gode grunde til at se DNA-replikationsprocessen som et produkt af intelligent design, og designtesen motiverer derfor én til at være skeptisk over for påstanden om, at følgestrengssyntesen er “kluntet” og dårligt designet. En skepsis, som en nærmere analyse af følgestrengssyntesen underbygger.
Til sammenligning er der intet i tesen om, at DNA-replikationsprocessen er et produkt af tilfældige mutationer og naturlig selektion, der motiverer én til at udvise skepsis over på påstanden om dårligt design. Hvis en struktur er dårligt designet, er det fordi den naturlige selektion har medført dette. Og hvis strukturen er godt designet, er det fordi den naturlige selektion har medført dette.
Hvad dette angår, er det således ID-kritikernes teori, der kan forklare alt og dermed intet forklarer.
Referencer
Prentø P. & Jensen P.V., 2000, Cellebiologi: Cellens organisation og livsprocesser, Gads Forlag.
Rensberger B., 1996, Life Itself: Exploring the Realm of the Living Cell, Oxford University Press.
Kategorier
