
Design og det første livs non-hierarkiske mønster

Forestil dig, at du skulle sammensætte et team til en mission på en fremmed og uudforsket planet. Teamet ville være uden radiokontakt til nogen andre og ville være nødt til at løse ethvert opstået problem udelukkende ved at bruge teammedlemmernes samlede færdigheder. Hvordan ville du sammensætte dit team?
Ville du udvælge en række personer, der alle havde præcis det samme snævre sæt af færdigheder, f.eks. et team af sprængningseksperter? Det ville næppe være en god idé, da teamet vil kunne komme ud for mange uforudsete problemer, inklusiv nogle, der kræver andre færdigheder end en sprængningseksperts.
Eller ville du udvælge en række personer, der havde et bredt sæt af de samme færdigheder, som de hver især mestrede middelmådigt? Det ville give teamet et bredere sæt af færdigheder, men der ville være en risiko for, at teamet kom ud for problemer, der krævede mere end en middelmådig beherskelse af en given færdighed.
Eller ville du udvælge en række personer, der hver især var ekspert inden for hver deres vidt forskellige felt? Altså en sprængningsekspert, en læge, en mekaniker, etc.? Med denne løsning ville du have det bedste af to verdener: Et bredt udvalg af færdigheder, som hver især kunne bruges på ekspertniveau.
De første designede livsformer
Hvis de første livsformer på Jorden var produktet af intelligent design (Gene, 2007; Krauze, 2019), ville designerne have stået over for en lignende udfordring. Hvordan skulle dette “team” af designede livsformer sammensættes, så det bedst var rustet til at løse de uforudsigelige situationer, som livet på den forhistoriske Jord kunne resultere i?
En løsning kunne være at designe en celle (eller en population af identiske celler), der løste sine grundlæggende biologiske opgaver – DNA-replikation, cellemembran-syntese, metabolisme, etc. – på én bestemt måde. Dette ville svare til at sammensætte et team udelukkende af sprængningseksperter, og ville gøre populationen af designede celler sårbare over for udfordringer, som deres snævre sæt af egenskaber ikke var egnet til at løse.
En anden løsning kunne være at designe en “supercelle” (eller en population af sådanne celler), med flere parallelle systemer og gener til at forfølge enhver metabolisk strategi. En sådan celle ville være i stand til at håndtere et langt større udsnit af udfordringer, men dette ville have sin pris: Cellen ville have et langt genom med gener for alle disse funktioner, og hver gang cellen skulle formere sig, ville den skulle kopiere alle disse gener.
En effektiv løsning ville være at designe en population af celler med vidt forskellige egenskaber og forskellige måder at løse deres grundlæggende biologiske opgaver, dvs. som en heterogen population. Og at give cellerne evnen til at “kommunikere” med hinanden ved at overføre gener til hinanden på tværs af arter. Så hvis en udfordring krævede en celle, der både havde systemerne “A” og “B”, ville en celle, der kun havde system A, kunne løse udfordringen, hvis den fik system B fra en anden celle.
Og meget tyder på, at det faktisk er den løsning, vi ser i eksisterende celler.
Manglende evolutionære forbindelser
Bakterier kopierer deres DNA med én slags replikationsmaskineri, mens arkæer kopierer deres med en anden slags replikationsmaskineri. De to mekanismer er så forskellige, at evolutionsbiologer anser DNA-replikationen i de to grupper som værende opstået uafhængigt af hinanden (Leipe et al., 1999).
Ligeledes er bakterier og arkæers cellemembraner opbygget af vidt forskellige fosfolipider (isoprenide glycerol-1-fosfat-ethere for arkæers vedkommende og glycerol-3-fosfat-fedtsyreestere for bakteriers vedkommende), syntetiseret af vidt forskellige enzymer – igen uden nogen tegn på evolutionær forbindelse mellem de to systemer (Koga et al., 1998).
Umiddelbart tyder det på, at den oprindelige population af designede celler i hvert fald har bestået af to forskellige celletyper, svarende til bakterier og arkæer. Og måske har hver af disse celletyper også været repræsenteret i flere forskellige varianter, således at der ikke blot har været én bakterie og én arkæ.
Hierarkiske mønstre og deres årsager
Når evolutionsbiologer skal argumentere for, at alle organismer er beslægtede, peger de ofte på tilstedeværelsen af et såkaldt indlejret hierarki, på engelsk nested hierarchy. Begrebet hentyder til den observation, at levende organismer kan klassificeres i et hierarkisk, grupper-i-grupper mønster. En mus kan således klassificeres som hørende til gruppen “pattedyr”, fordi den har pels. Gruppen af pattedyr kan igen klassificeres som hørende til gruppen “amnioter”, fordi deres fostre udvikler sig i en fosterhinde, etc. (figur 1)
Argumentet for fælles afstamning er, at hver af de træk, man bruger til at klassificere grupperne med (pels, fosterhinde, etc.) var til stede i denne gruppes sidste fælles stamform, hvorfra trækket er blevet nedarvet af gruppens medlemmer.
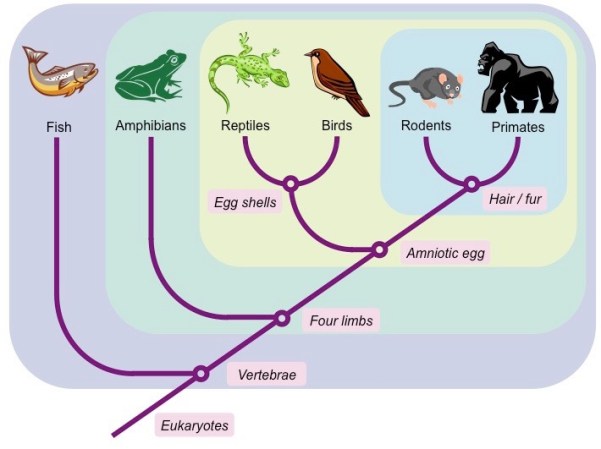
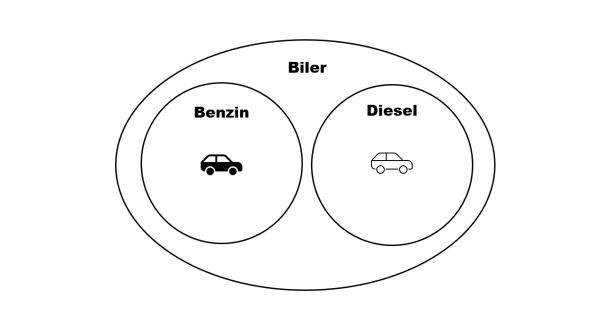
Hierarkiske mønstre kan i nogen grad også observeres i designede strukturer, der ikke er beslægtede med hinanden i nogen biologisk forstand. Dette skyldes, at indledende designbeslutninger kan indskrænke det efterfølgende felt af mulige designbeslutninger. Beslutter man f.eks. at designe en benzinbil, har man dermed også forpligtet sig til at indrette den med en række komponenter – cylindre, brændstoftilførsel, etc. – der er velegnet til forbrændingen af benzin, men som ikke er velegnet til forbrænding af diesel. Dermed kan benzinbiler “klassificeres” ud fra en række fællestræk, der adskiller sig fra dieselbilers fællestræk (figur 2).
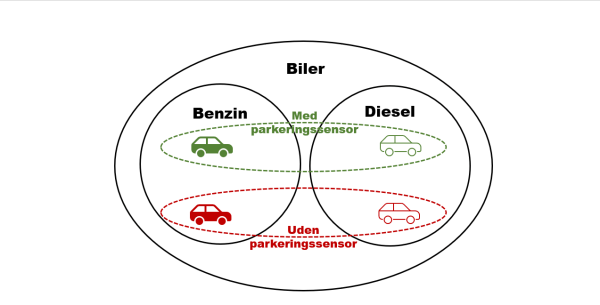
Designede objekter er dog i høj grad karakteriseret ved non-hierarkiske mønstre. Dette skyldes, at hvor indledende designbeslutninger ikke indskrænker det efterfølgende felt, er en designer fri til at genbruge et design i forskellige modeller. F.eks. kan en bil udstyres med parkeringssensor, uanset om den er en benzin- eller dieselbil. Dermed opstår grupperinger, der går på tværs af hinanden (figur 3).
Livets non-hierarkiske mønster
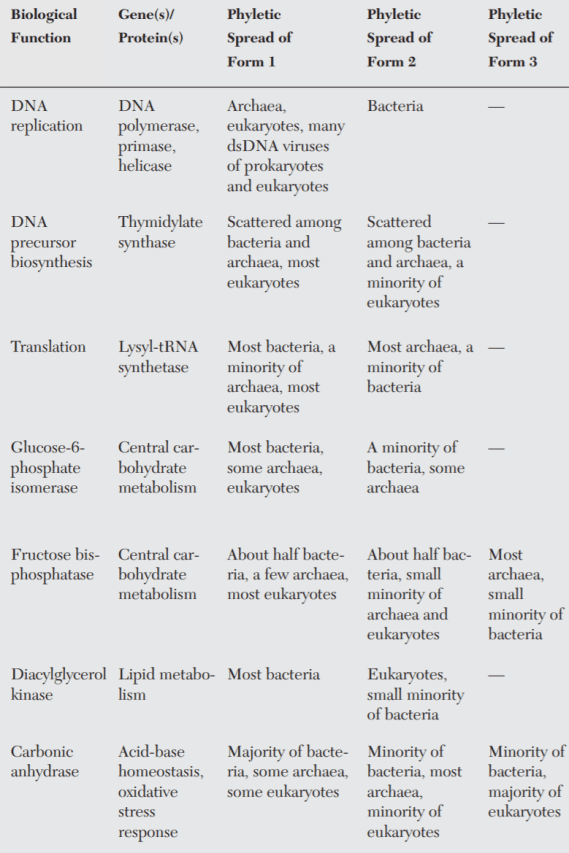
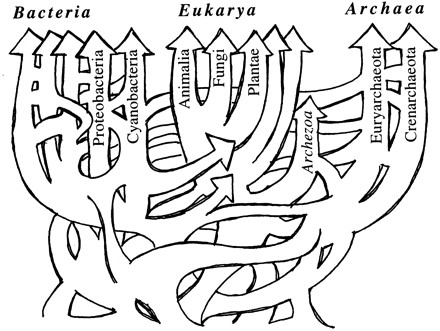
Når vi retter blikket mod mikrober – der er de livsformer, der er tættest på livets oprindelse – ser vi, at de i høj grad er opbygget efter et non-hierarkisk mønster. Gener og proteiner for centrale og informationshåndterende og metaboliske processer i cellen viser sig således at være fordelt på kryds og tværs af bakterier og arkæer (figur 4).
Dette non-hierarkiske mønster er en af årsagerne til, at nogle forskere har opgivet tanken om, at alt liv nedstammer fra en enkelt sidste fælles stamform – på engelsk Last Universal Common Ancestor, eller blot LUCA.
“LUCA er en dybt problematisk størrelse,” skriver mikrobiologen Ford Doolittle, hvorefter han tilføjer: “Faktisk er LUCA en illusion.” (Doolittle, 2009, s. 2224). Han fik støtte af den afdøde Carl Woese, hvis arbejde med fylogenier baseret på 16S rRNA gjorde ham berømt:”Den universelle stamform er ikke en størrelse, ikke en ting. Det er en proces, der er karakteristisk for et specifikt evolutionært stadie.” (Woese, 1998, s. 6858). I en senere artikel var han endnu mere direkte: “Der har aldrig været nogen universel fælles stamform. Doktrinen om Fælles Afstamning har snydt os.” (Woese, 2005, s. 110)
Forskere som Woese og Doolittle mener, at den tidligste evolution var præget af en høj grad af horisontal genoverførsel, og hvor gener således blev overført til arter, hvor vi ifølge tesen om universel fælles afstamning ikke ville forvente at finde dem. Denne genoverførsel var så omfattende, at det ikke giver mening at tale om “livets træ”, men snarere om “livets tjørnekrat” (figur 5).
Konklusion
Hvis de første livsformer var produktet af intelligent design, ville der være gode grunde til at designe dem som en heterogen population af celler med en non-hierarkisk fordeling af centrale gener/proteiner. Jeg har i denne artikel gennemgået observationer, der understøtter et scenario, hvori denne oprindelige designede population bestod af mindst to forskellige celletyper, svarende til bakterier og arkæer, der muligvis har været repræsenteret i forskellige varianter.
Referencer
Gene M., 2007, The Design Matrix: A Consilience of Clues, Arbor Vitae Press.
Koga Y., Kyuragi T., Nishihara M., Sone N., 1998, “Did archaeal and bacterial cells arise independently from noncellular precursors? A hypothesis stating that the advent of membrane phospholipid with enantiomeric glycerophosphate backbones caused the separation of the two lines of descent”, Journal of Molecular Evolution 46(1):54-63. https://doi.org/10.1007/pl00006283
Koonin E.V., 2011, The Logic of Chance: The Nature and Origin of Biological Evolution, FT Press.
Krauze M., 2019, “Intelligent design, information og livets oprindelse”, Facebook-note. https://www.facebook.com/notes/mikael-krauze/intelligent-design-information-og-livets-oprindelse/121359529187099/
Leipe D.D., Aravind L. & Koonin, E.V., 1999, “Did DNA replication evolve twice independently?”, Nucleic Acids Research 27(17):3389-401. https://doi.org/10.1093/nar/27.17.3389
Doolittle W.F., 1999, “Phylogenetic Classification and the Universal Tree”, Science 284(5423):2124-8. https://doi.org/10.1126/science.284.5423.2124
Doolittle W.F., 2009, “The practice of classification and the theory of evolution, and what the demise of Charles Darwin’s tree of life hypothesis means for both of them”, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 364(1527):2221-8. https://doi.org/10.1098/rstb.2009.0032
Woese C., 1998, “The universal ancestor”, Proceedings of the National Academy of Sciences of the United States of America 95(12):6854-9. https://doi.org/10.1073/pnas.95.12.6854
Woese C., 2005, “Seeing the Present”, i Sapp J. (red.), Microbial Phylogeny and Evolution: Concepts and Controversies, Oxford University Press.
Kategorier
