
Fælles design og hexameriske helicaser
Jeg har tidligere argumenteret for intelligent design (ID) af livets oprindelse som noget, en rationel person kan overveje som en reel mulighed. (Krauze, 2019). Dette argument er foreneligt med, men nødvendiggør ikke, at de oprindeligt designede celler var identiske, og at alle nulevende organismer således nedstammer fra, hvad der i princippet er en enkelt stamform. Argumentet er også foreneligt med, at de oprindeligt designede celler var forskellige, og at det således ikke lader sig gøre at føre alle nulevende organismer tilbage til en enkelt stamform, hvorfra de alle nedstammer. Argumentet lader os således ikke skelne mellem, om livet er monofyletisk (én oprindelse) eller polyfyletisk (flere oprindelser).
Det vil jeg råde bod på i denne artikel, hvor jeg vil beskrive begrebet “fælles design” og vise, hvorledes det kan kaste lys over livets udvikling. Jeg vil illustrere begrebet gennem en diskussion af hexameriske helicaser, der spiller en central rolle i DNA-replikation, og argumentere for, at det inden for et design-paradigme er en rationel mulighed, at det første liv bestod af mindst to ubeslægtede celletyper, og at livet således er polyfyletisk.
Fælles afstamning og konvergens
Lige siden Aristoteles har det stået klart, at forskellige organismer har bestemte træk og strukturer til fælles. Alle fugle har fjer, alle pattedyr producerer mælk og så videre. Inden for traditionel evolutionsbiologi kan strukturer, der ligner hinanden, dybest set forklares på to forskellige måder: Som et eksempel på fælles afstamning (på engelsk, common descent) eller som et eksempel på konvergens (convergence).
Ligheder, der skyldes fælles afstamning, kaldes homologier (Futuyma, 1998). Hvis to arter begge besidder en homolog struktur, menes det således, at denne struktur er nedarvet fra en fælles forfader, der også besad strukturen. Fjerene på en svale og på en pingvin betegnes således som homologe, da begge arter menes at have arvet det at have fjer fra en fælles forfader til alle fugle.
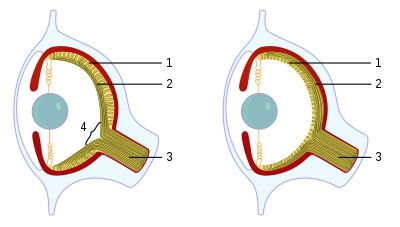
Ligheder, der skyldes konvergens, kaldes homoplasier (Futuyma, 1998). Dette begreb betegner ligheder, der ikke er nedarvet fra en fælles forfader, men har udviklet sig uafhængigt af hinanden. Et eksempel er øjnene hos hvirveldyr og blæksprutter (Cephalopoda), der har stor lighed med hinanden (figur 1), selv om den fælles forfader til blæksprutter og hvirveldyr ikke menes at have haft en sådan øjetype.
Fælles design
Som beskrevet i den foregående sektion, har traditionel evolutionsbiologi to “værktøjer” til at forklare ligheder. Et design-paradigme har også adgang til disse værktøjer, da en oprindeligt designet livsform kan udvikle sig og således give ophav til en række arter, der både er i besiddelse af homologier og homplasier.
Et design-paradigme har imidlertid et tredje værktøj i værktøjskassen; fælles design (common design), altså at ligheder kan skyldes, at strukturerne er designet for at udføre den samme funktion. Ræsonnementet bag fælles design er ingeniørprincippet om at lade form følge funktion. Og det er vores erfaring med intelligente designere, at de ofte genbruger løsninger, når de står over for lignende problemer.
Vi kan skelne mellem to grundlæggende typer af fælles design:
Genbrug af struktur: At genbruge en struktur med få eller ingen ændringer til at løse det samme problem igen og igen. Dette kender vi fra IKEA-møbler, der samles med 12 identiske skruer.
Genbrug af idé: At genbruge en overordnet idé til at løse det samme problem i en ny kontekst. Resultatet er to strukturer, der umiddelbart kan ligne hinanden, men som er fundamentalt forskellige.
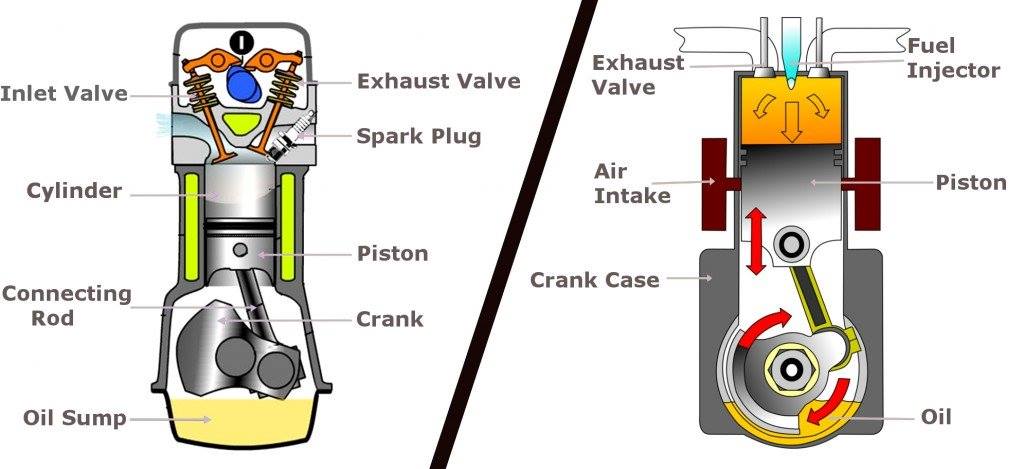
Princippet om genbrug af idé kan illustreres ved forskellen mellem cylinderne i en benzinbil og en dieselbil (figur 2). De to cylindertyper ligner umiddelbart hinanden, da de begge forsøger at løse det samme designproblem – at omdanne ekspansionsenergien fra forbrændingen af et brændstof til en cirkulær bevægelse. Men på grund af de kemiske forskelle mellem benzin og diesel er de to cylindertyper nødt til at løse problemet på to forskellige måder og er således fundamentalt forskellige.
Fælles design er således et begreb, som er velkendt fra menneskeskabte objekter. Men kan begrebet også give mening i en biologisk kontekst?
Hexameriske helicaser og DNA-replikation
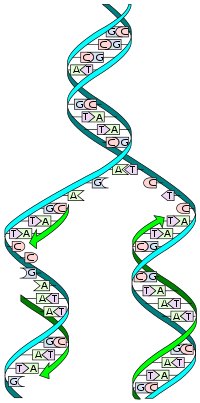
Cellens arvemateriale er DNA-molekylet, en dobbelthelix med genetiske instruktioner kodet langs de to komplementære strenge. Ved replikation af DNA opsplittes de to strenge og hver streng bruges som skabelon for replikation af en ny komplementærstreng (figur 3).
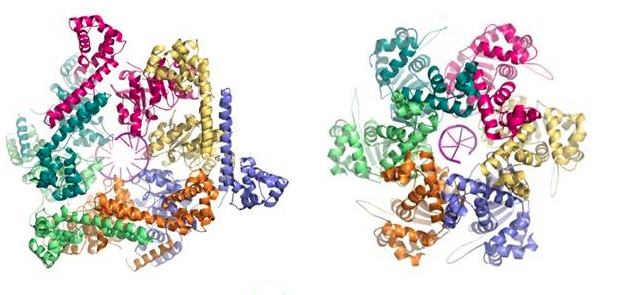
Opsplitningen af DNA-molekylet udføres af et protein kaldet helicase (figur 4). Replikations-helicasen er en ringformet hexamer, dvs. det er en ring bestående af seks identiske molekyler. Hver af disse molekyler er håndvægtsformede, hvilket får helicasen til at ligne to baderinge stablet oven på hinanden. Helicasen menes at fungere ved at omslutte den ene DNA-streng og adskille den fra den anden i en process, der kaldes “sterisk eksklusion” (O’Donnell & Li, 2018).
Fælles afstemning som forklaring?
Alle celler bruger DNA som arvemateriale, og alle celler bruger ringformede, hexameriske helicaser til at opsplitte DNA-strengene som led i replikationen. Dette gælder både bakterier, eukaryoter (celler med en cellekerne, der omfatter både dyr, planter og svampe) og arkæer (en celletype, der blandt andet findes i ekstremt varme miljø som hydrotermiske gejsere).
Så det må være ret ligetil at forklare dette gennem fælles afstamning, hvor alle nulevende celler har arvet deres helicaser fra en fælles stamfader, hvor de udførte samme funktion?
Næppe. I en fagfællebedømt artikel med den provokerende titel “Did DNA replication evolve twice independently?” argumenterer tre evolutionsbiologer for, at replikationsmaskineriet i bakterier på den ene side og eukaryoter og arkæer på den anden side er så forskellig fra hinanden, at de to mekanismer er opstået uafhængigt af hinanden (Leipe et al., 1999).
Den nu afdøde mikrobiolog og National Academy of Sciences-medlem Carl Woese delte denne opfattelse: “Så godt som ingen homologi eksisterer mellem den bakterielle genom-replikationsmekanisme og den, som arkæer og eukaryoter har til fælles. … Moderne genom-replikationsmekanismer ser ud til at være udviklet to gange.” (Woese, 2002, s. 8743)
Et nyligt review af replikations-helicaser formulerer det direkte: “Der er ingen evolutionær sammenhæng mellem de hexameriske helicaser fra bakterier og arkæer/eukaryoter” (O’Donnell & Huilin, 2018, s. 122).
Denne konklusion er blandt andet baseret på opdagelsen af, at helicasen ikke binder sig til den samme DNA-streng i bakterier og eukaryoter, og at den heller ikke vender samme vej (figur 5). At forklare oprindelsen af en type helicase gennem den gradvise ændring af en anden type, samtidig med at den udfører sin opgave i DNA-replikationen, er således ikke tiltrækkende.
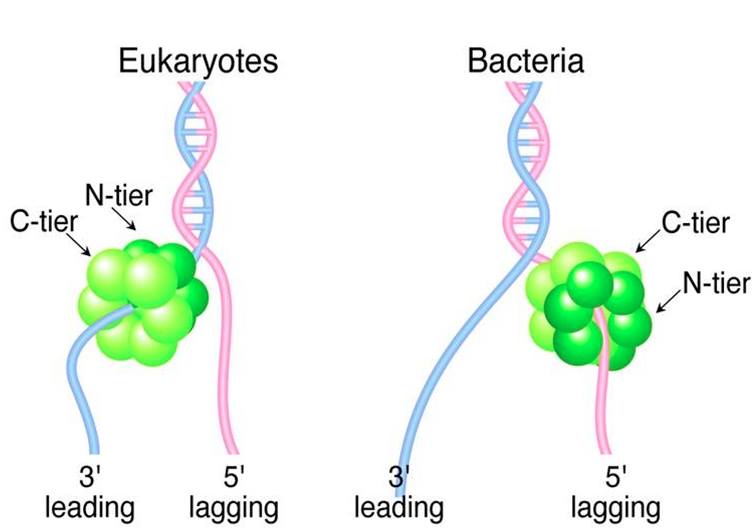
Konvergens som forklaring?
Som vi har set, er fælles afstamning ikke en sandsynlig forklaring på lighederne mellem den hexameriske helicase i henholdsvis bakterier og eukaryoter. Hvad med det andet værktøj i den forklaringsmæssige værktøjskasse – konvergens?
Hvis de hexameriske helicaser er opstået gennem konvergent evolution, må der engang have været et tidspunkt, hvor alle livsformer på Jorden replikerede deres DNA uden brug af helicaser. Fra denne helicase-løse tilstand må de to forskellige helicase-typer være opstået uafhængigt af hinanden.
Der er imidlertid grund til at være skeptisk over for dette. Brugen af helicaser ved DNA-replikation er universel; alle celler bruger enten den ene eller den anden type. Vi kender ikke til nogen celler, der kopierer deres DNA uden brug af en helicase. Hvis der engang har eksisteret celler, der kopierede deres DNA uden brug af en helicase, har de ikke efterladt sig nogen efterkommere, der har bevaret dette træk.
Nogle evolutionsbiologer peger på vira, der ikke har helicaser, og argumenterer for, at disse kan repræsentere en tidlig, helicase-løs fortid (Yao & O’Donnell, 2016).
Det skal dog huskes på, at vira er intracellulære parasitter. Modsat celler kan de ikke selvreplikere, men er nødt til at kapre cellens maskineri og bruge dette til at blive replikeret. Dette gør det vanskeligt at forestille sig en tid, hvor livet på Jorden udelukkende bestod af vira i deres nuværende form.
“Fordi de er simple, blev vira omfavnet af mange biologer som det manglende led mellem liv og ikke-liv,” konstaterer Moreira og López-García i fagtidsskriftet Nature Reviews Microbiology. Som de to forfattere pointerer, er regressiv evolution en fundamental proces for parasitter; de mister gener og bliver simplere, fordi mange af deres processer udføres af værtsorganismen. “Således er simple vira ikke tegn på, at vira er gamle [i en evolutionær forstand] eller at de er primitive, men er en konsekvens af parasitisme.” (Moreira & López-García, 2009, s. 5)
Med andre ord: Hvis vira er beslægtede med celler, er der snarere tale om, at vira er opstået ved reduktion fra en celle-lignende tilstand end omvendt.
Der er således ingen evidens for, at nulevende organismer nedstammer fra organismer, der formerede sig uden brug af helicaser. Og dermed heller ingen evidens for, at dette træk har udviklet sig konvergent.
Fælles design som forklaring
I et design-paradigme er man ikke begrænset til at skulle vælge mellem fælles afstamning og konvergens. En tredje mulighed står åben, nemlig fælles design.
En “fælles design”-analyse af de hexameriske helicaser i henholdsvis bakterier og eukaryoter starter med at konstatere, at de to strukturer begge løser det samme grundlæggende design-problem: Hvordan adskiller man det dobbeltstrengede DNA, så replikation bliver mulig? Denne opgave løses gennem en ringformet struktur, der slutter sig om DNA-strengen og sikrer, at helicasen følger denne, mens replikationen foregår.
Men hvis funktionen er den samme, hvad forklarer da den store forskel på de to helicaser? Et svar lader til at komme os i møde ved at se helicaserne som et eksempel på genbrug af en idé, som beskrevet ovenfor i eksemplet med benzin- og dieselcylindre. I denne optik er de forskellige helicaser to måder at løse det samme problem i forskellige kontekster, henholdsvis den bakterielle og den eukaryote celle. Resultatet er to strukturer, der legemliggør den samme design-logik på to vidt forskellige måder. Og som på trods af deres umiddelbare ligheder derfor er fundamentalt forskellige.
Bakterier og eukaryoter er to fundamentalt forskellige celletyper. En af forskellene består i længden på deres genom, hvor en eukaryots genom i gennemsnit er markant længere end en bakteries genom – sammenlign bakterien E. coli med dens 4 mio. basepar med den eukaryote gærcelle S. cerevisiae med dens 14 mio. basepar. I bakterier er genomet organiseret som et enkelt cirkulært kromosom, mens det i eukaryoter er organiseret i et antal tætpakkede kromosomer.
Lange DNA-strenge har en tendens til at filtre sig sammen, ikke ulig et par iPhone-høretelefoner, man har i lommen. For at forhindre, at DNA-strengen bliver filtret, organiseres den i den eukaryote celle i nukleosomer, hvor DNA’et pakkes omkring kugleformede proteiner kaldet histoner (figur 6).
Arkæer har ligesom bakterier korte genomer. Ikke desto mindre er arkæers DNA også pakket i nukleosomer omkring histoner (Bailey et al., 2002; Henneman et al., 2018).
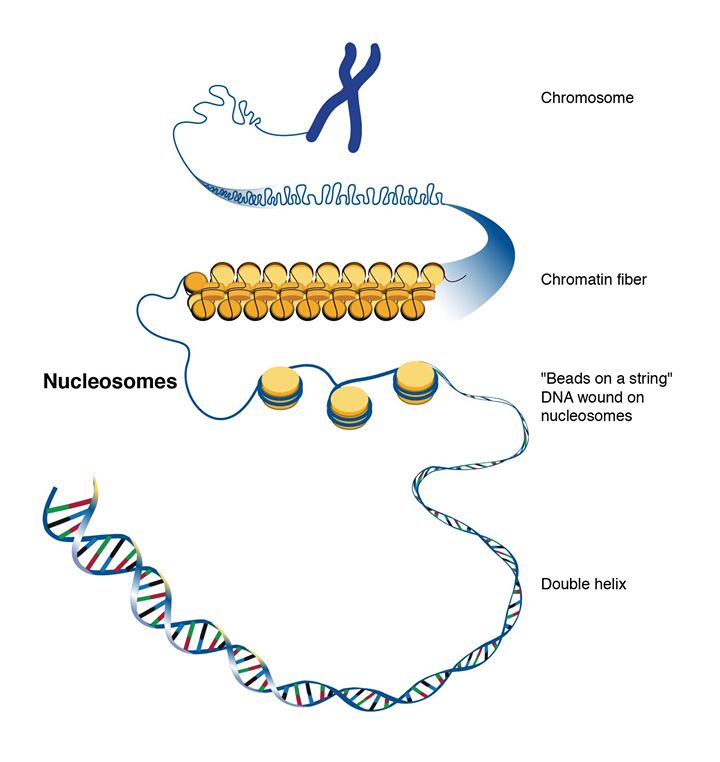
Studier tyder på, at helicaser spiller en rolle, når DNA-strengen skal vikles ud af nukleosomet (Eggleston et al., 1995; Brennan et al., 2016). Eukaryoter og arkæers helicasers funktion er således ikke blot at adskille det dobbeltstrengede DNA, men også at adskille DNA-strengen fra nukleosomet.
Der er endnu mange aspekter af helicasens funktion, man ikke kender til. Men ud fra en “fælles design”-analyse kan vi fremsætte en forudsigelse, der kan efterprøves: De aspekter af den eukaryote/arkæale helicase, der gør den i stand til at adskille DNA-strengen fra nukleosomet, vil vise sig også at være de aspekter, der gør den forskellig fra den bakterielle helicase. Med andre ord, forskellene mellem de to typer helicaser vil vise sig at kunne forklares med henvisning til, at de to typer helicaser udfører den samme funktion i forskellige cellulære kontekster.
Konklusion
Jeg har i det ovenstående beskrevet begrebet “fælles design” og vist, hvordan det kan bruges til at analysere biologiske strukturer og dermed kaste lys over livets udvikling. Det er således et begreb, der under et design-paradigme bør høre til i den forklaringsmæssige værktøjskasse sammen med fælles afstamning og konvergens.
De hexameriske replikations-helicaser lader til at være et godt eksempel på fælles design. Helicaserne fra henholdsvis bakterier og eukaryoter/arkæer udviser fundamentale forskelle, der gør det svært at argumentere for, at de to typer helicase er evolutionært beslægtede med hinanden. Samtidig er der ingen evidens for, at nulevende organismer nedstammer fra organismer, der formerede sig uden brug af helicaser. Og dermed er der heller ingen evidens for, at helicaserne er opstået gennem konvergent evolution.
Fælles design er den eneste forklaring, der kan redegøre for både de overordnede ligheder og de fundamentale forskelle, der eksisterer mellem de hexameriske replikations-helicaser. Fælles design er en hypotese, der fremsætter forudsigelser, der kan efterprøves, og som derfor kan anspore fremtidig forskning.
Opdateret 9. april 2020 med diskussion af vira.
Referencer
Bailey K.A., Marc F., Sandman K., Reeve J.N., 2002, “Both DNA and histone fold sequences contribute to archaeal nucleosome stability”, Journal of Biological Chemistry 277(11):9293-301. https://doi.org/10.1074/jbc.M110029200
Brennan L., Forties R., Patel S., et al., 2016, “DNA looping mediates nucleosome transfer”, Nature Communications 7:13337. https://doi.org/10.1038/ncomms13337
Eggleston A.K., O’Neill T.E., Bradbury E.M. & Kowalczykowski S.C., 1995, “Unwinding of nucleosomal DNA by a DNA helicase”, Journal of Biological Chemistry 270:2024-31. https://doi.org/10.1074/jbc.270.5.2024
Futuyma D.J., 1998, Evolutionary Biology, Third Edition, Sinauer. ISBN 0-87893-189-9
Henneman B., van Emmerik C., van Ingen H., Dame R.T., 2018, “Structure and function of archaeal histones”, PLoS Genetics 14(9):e1007582. https://doi.org/10.1371/journal.pgen.1007582
Krauze M., 2019, “Intelligent design, information og livets oprindelse”, Facebook-note. https://www.facebook.com/notes/mikael-krauze/intelligent-design-information-og-livets-oprindelse/121359529187099/
Leipe D.D., Aravind L. & Koonin, E.V., 1999, “Did DNA replication evolve twice independently?”, Nucleic Acids Research 27(17):3389-401. https://doi.org/10.1093/nar/27.17.3389
Moreira D. & López-García P., 2009, “Ten reasons to exclude viruses from the tree of life”, Nature Reviews Microbiology 7:306-11. https://doi.org/10.1038/nrmicro2108
O’Donnell M. E. & Li H., 2018, “The ring-shaped hexameric helicases that function at DNA replication forks”, Nature Structural & Molecular Biology 25(2):122-30. https://doi.org/10.1038/s41594-018-0024-x
Woese C.R., 2002, “On the evolution of cells”, PNAS 99(13)8742-7. https://doi.org/10.1073/pnas.132266999
Yao N.Y. & O’Donnell M.E., 2016, “Evolution of replication machines”, Critical Reviews in Biochemistry and Molecular Biology 51(3):135-49. https://doi.org/10.3109/10409238.2015.1125845
Kategorier

Hej Martin Krauze
Tak for denne spændende gennemgang. Jeg har et sted set arkæer omtalt som værende tilstede også i det menneskelige mikrobiom, men kan ikke finde det bekræftet. Har du nogen kommentarer til dette?
Mvh Steffen Kjærulff
Hej Steffen,
Tak for dit spørgsmål. Det er korrekt forstået, at der foruden bakterier og vira også er arkæer til stede i det menneskelige mikrobiom, altså det økosystem af mikroorganismer, der findes på og i vores krop. Nedenstående open access review går mere i dybden med dette:
Lurie-Weinberger M.N., Gophna U., 2015, “Archaea in and on the Human Body: Health Implications and Future Directions”, PLoS Pathogens 11(6): e1004833. https://doi.org/10.1371/journal.ppat.1004833