
Den bakterielle flagel: En irreducibelt kompleks maskine
Introduktion
Den bakterielle flagel er en af naturens mest fantastiske og velundersøgte nanomaskiner (Chen et al. 2011). Karakteriseret som “naturens mest kraftfulde roterende motor” (Pallen et al. 2005, s. 147) består den af en proptrækkerformet propel, drevet af en transmembran ion/proton-strøm, selvkonstrueret ved hjælp af et sindrigt sekretionssystem. En artikel i tidsskriftet New Scientist beskriver flagellen som “en af mest komplekse og elegante stykker biologisk maskineri, som vi kender … en kompliceret nanomaskine, som ingen menneskelig ingeniør ville kunne eftergøre” (Jones 2008, s. 41).
Flagellen spiller en central rolle i biokemikeren Michael Behes bog, Darwin’s Black Box: The Biochemical Challenge to Evolution ( 1996), hvor Behe karakteriserer flagellen som et irreducibelt komplekst system.
Behe definerer et irreducibelt komplekst system som “et system sammensat af adskillige samvirkende og til hinanden tilpassede dele, der bidrager til den grundlæggende funktion, og hvor fjernelsen af en hvilken som helst af disse dele vil få systemet til at ophøre med at fungere” (Behe 1996, s. 39).
Et irreducibelt komplekst system, argumenterer Behe, “kan ikke produceres direkte (det vil sige, ved løbende at forbedre den oprindelige funktion, som fortsætter med at virke gennem den samme mekanisme) ved små på hinanden følgende ændringer af et forløber-system” (Behe 1996, s. 39).
Det er nu 25 år siden, Behes bog udkom. I mellemtiden er vores viden om den bakterielle flagel blevet kraftigt forøget, og vi har desuden fået større kendskab til de mange varianter af flagellen, der eksisterer. Flageller er fundet i fjernt beslægtede bakterier, og der er stor varians i, hvor mange komponenter de forskellige flageller består af (Pallen et al. 2005). Dog er der også en konserveret kerne af komponenter, som findes i alle undersøgte flageller (Chen et al. 2011).
I denne artikel vil jeg give en detaljeret beskrivelse af den bakterielle flagel, opdateret med den viden, der er kommet til siden Behes bog. På baggrund af den konserverede kerne af komponenter og fylogenetiske analyser af flagelbærende bakterier vil jeg rekonstruere “ur-flagellen”, som den har været til stede i den sidste fælles stamform til alle flagelbærende bakterier. Jeg vil kombinere denne analyse med data fra såkaldte “knock-out” eksperimenter og identificere ur-flagellens irreducible kerne.
Den irreducible kerne er det sæt af dele, der alle er nødvendige for at et givent system kan udføre sin funktion (Thomas 1999; Dembski 2002). Logikken er, at hvis en del mangler i en naturligt forekommende bakterie med en flagel, eller hvis man i et laboratorium kan fjerne genet uden at fjerne flagellens funktion, er det pågældende gen ikke del af den irreducible kerne.
Flagellens opbygning og konstruktion
Et billede siger mere end tusind ord; denne korte video (5:13) om flagellens opbygning og konstruktion kan anbefales som supplement til nedenstående beskrivelse.
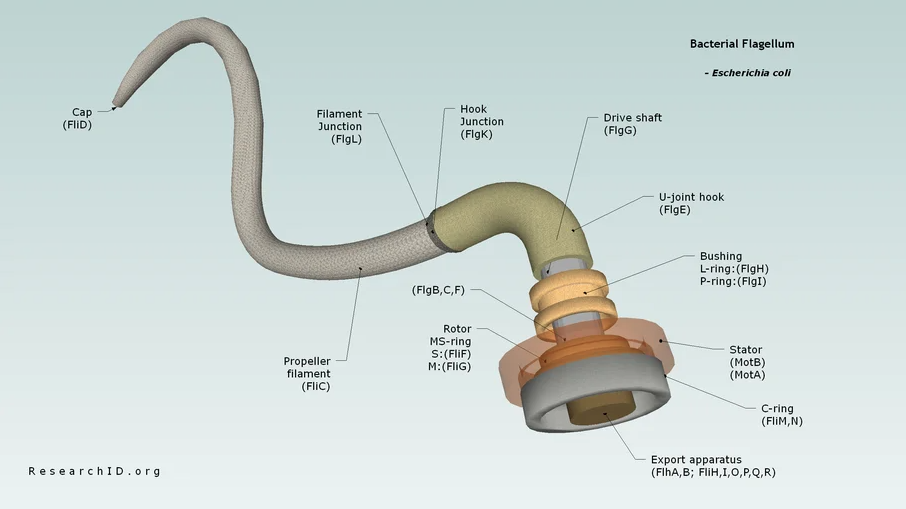
Den bakterielle flagel (figur 1) er en roterende motor med en piskelignende svingtråd, som gør bakterier i stand til at bevæge sig gennem vand og andre flydende miljøer (DeRosier 1998; Diepold & Armitage 2015; Nakamura & Minamino 2019).
Den bakterielle flagel skal ikke forveksles med den eukaryote flagel (også kaldet ciliet eller undulipodiet) eller den arkæale flagel (også kaldet arkellen), der er opstået uafhængigt af den bakterielle flagel. I det følgende vil jeg for nemheds skyld blot omtale den bakterielle flagel som “flagellen”; den eukaryote og den arkæaele flagel vil ikke blive omtalt yderligere.
Opbygning
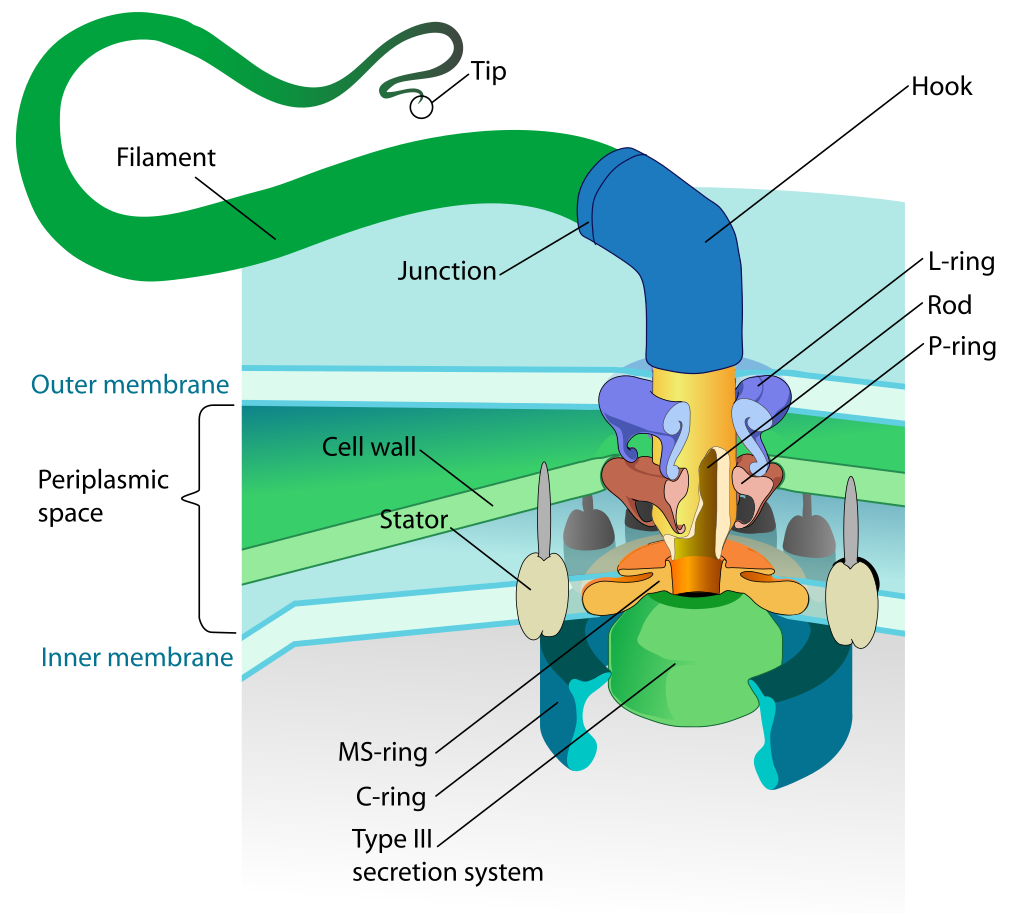
Flagellen består af en motor, en svingtråd, en aksel og en krog, samt en række ringe og et sekretionssystem.
Flagellens motor, der er forankret i cellemembranen, er drevet af en indstrømning af protoner eller natrium-ioner. Svingtråden befinder sig uden for cellen, hvor dens roterende bevægelse genererer fremdrift, som skruen på et skib.
Den roterende bevægelse genereres i det såkaldte motor/stator-kompleks, der er forankret i peptidoglykanlaget og som en dynamo omdanner kraften fra indstrømningen af protoner eller ioner til en roterende bevægelse (Santiveri et al. 2020). Den roterende bevægelse overføres til rotor-komplekset, eller C-ringen, hvorfra den overføres gennem akslen. Herfra overføres den roterende bevægelse gennem krogen, der fungerer som kardanaksel, hvorefter den overføres til svingtråden. MS-ringen forankrer flagellen i den indre cellemembran og er således ikke en roterende del.
Hos de fleste arter kan flagellen rotere både med og mod uret. I E. coli, hvor flagellen er mest grundigt studeret, forårsager rotation mod uret en fremadrettet bevægelse af bakterien (i fagsprog, at bakterien “løber”), mens rotation med uret forårsager en reorientering af bakterien i en tilfældig retning (bakterien “tumler”).
Flagellens rotationsretning reguleres af det kemotaksiske system. Dette system er strengt taget ikke nødvendigt for flagellens evne til at udføre sin funktion, på samme måde som en GPS ikke er nødvendig for en bils evne til at køre. En flagel uden kemotaksisk system kan f.eks. bruges af bakterien til at fjerne sig fra et overfyldt miljø ved tilfældig spredning. Det kemotaksiske system er fraværende i den flagelbærende bakterie Aquifex aeolicus, der dog muligvis har et termotaksisk (dvs. temperaturstyret) system i stedet (Deckert et al. 1998).
Konstruktion
Med en længde på 5-20 μm er flagellen en forholdsvis lang struktur, op til flere gange bakteriens egen længde. Konstruktionen af en struktur på den længde udgør et designproblem, da strukturen ikke kan konstrueres inde i cellen, men må konstrueres in situ uden for cellemembranen.
Flagellen konstrueres indefra og ud, ved hjælp af et sekretionssystem, der transporterer materialer ud gennem de dele af flagellen, der allerede er konstrueret. Man kan forestille sig en skorsten, der bygges ved at man hejser mursten op inde i skorstenen, mens den bygges.
Først samles MS-ringen i cellemembranen, muligvis sammen med dele af sekretionssystemet. Dernæst samles motor/stator-komplekset rundt om MS-ringen, efterfulgt af C-ringen. Disse dele udgør det fundament, fra hvilken resten af flagellen konstrueres (Macnab 2003).
Dernæst samles akslen, efterfulgt af L- og P-ringene (i gram-negative arter), krogen og svingtråd/krog-samlingen. Dernæst samles svingtråden (FliC) med hjælp fra svingtrådskapslen (FliD). Svingtrådskapslen er ikke blot en passiv “prop”, men faciliterer foldningen og placeringen af FliC (Macnab 2003).
Flagellens grundlæggende funktion
Behes definition af irreducibel kompleksitet henviser til et systems “grundlæggende funktion”. I en biologisk kontekst kan et systems funktion defineres som den egenskab ved systemet, der bidrager til organismens fitness (Thain & Hickman 2000, s. 252; se også Ruse 2003 og Perlman 2009).
Spørgsmålet om, hvad der udgør et systems grundlæggende funktion, er mere kompliceret, da et system kan have flere funktioner. For eksempel bruger fugle deres fjer til at flyve med, men de bruger dem også til varmeisolering (hvilket muligvis er deres oprindelige funktion, se Bock 2000). Hvilken af disse er fjerenes grundlæggende funktion?
For at svare på dette, må vi se på, hvilke af systemets egenskaber, der bedst forklares gennem deres bidrag til de respektive funktioner. Tag en fugls svingfjer: De er asymetriske og deres stråler er forsynet med små kroger, der griber fat i nabostrålen og giver fjeren en glat overflade. Fjer uden disse egenskaber (som de dun, der dækker nyudklækkede fugleunger) kan også bruges til varmeisolering, men de kan ikke bruges til at flyve med. Svingfjerens evne til at gøre fugle i stand til at flyve forklarer således flere af fjerens egenskaber, og dette må derfor vurderes som dens grundlæggende, eller primære funktion.
Ligeledes er den bakterielle flagels grundlæggende, eller primære funktion at gøre bakterien i stand til at bevæge sig. Flagellen har også andre funktioner, såsom via svingtråden at binde til celler og andre overflader (Moens & Vanderleyden 1996). Men når vi kigger på alle flagellens egenskaber – ikke blot svingtrådens bindingsevne, men også dens evne til at antage en proptrækkerform, flagellens evne til at rotere, tilstedeværelsen af rotor, stator, etc. – står det klart, at de fleste egenskaber bedst forklares gennem deres bidrag til bakteriens evne til at bevæge sig.
Flagellen og fælles afstamning
Konventionel evolutionsbiologi er baseret på tesen om universel fælles afstamning, ifølge hvilken alle nulevende organismer nedstammer fra en enkelt fælles stamform (Sober & Steel, 2002). Det følger heraf, at alle flagelbærende bakterier nedstammer fra en fælles, flagelbærende stamform, der har været i besiddelse af den såkaldte “ur-flagel”.
Ud fra et designperspektiv kan vi forholde os mere skeptiske til tesen om universel fælles afstamning. Jeg har tidligere skrevet om muligheden af, at bakterier og arkæer ikke har en fælles stamform (Krauze 2020a), og at alle bakterier (og alle arkæer) muligvis heller ikke har det (Krauze 2020b).
Hvis man plotter flagelbærende bakterier ind på et evolutionært stamtræ, er det svært at få resultatet til at passe med, at flagellen er opstået i én art, hvorfra den er blevet nedarvet til alle nulevende flagelbærende arter (Snyder et al. 2009). Inden for et evolutionært paradigme tolkes dette som tegn på, at flere grupper har mistet deres flageller uafhængigt af hinanden, og/eller at gener for flagellen er blevet overført mellem arter. Men fra et designperspektiv kan det også ses som tegn på, at nogle af grupperne af flagelbærende bakterier ikke nedstammer fra en fælles stamform.
Ikke desto mindre vil jeg i det nedenstående tage udgangspunkt i konventionelle bakteriefylogenier og gå ud fra, at alle flagelbærende bakterier nedstammer fra en fælles stamform. Hvis min analyse skal kunne indgå i en diskussion om, hvorvidt flagellen er et produktet af en evolutionær proces, må analysen være baseret på antagelser, som også en tilhænger af konventionel evolutionsbiologi kan acceptere.
Ur-flagellen og dens irreducible kerne
Omkring 50 forskellige gener er involveret i konstruktionen og funktionen af den bakterielle flagel. Af disse 50 flagelrelaterede gener koder de 20-30 (afhængig af bakteriens art) for de genprodukter, der udgør selve flagellen. Omkring 10 gener er regulatoriske gener, der regulerer, hvornår andre gener udtrykkes, og omkring 10 gener er nødvendige for det ovenfor beskrevne kemotaksiske system (DeRosier 1998).
I min analyse vil jeg kun beskæftige mig med de gener, der udgør selve flagellen, og ikke beskæftige mig med de regulatoriske gener eller det kemotaksiske system. Dette er et bevidst valg, da jeg ikke ønsker at komme til at overdrive flagellens kompleksitet; om noget er min analyse en underdrivelse af, hvor kompleks flagellen er.
For at identificere ur-flagellens gener har jeg udvalgt 15 flagelbærende bakteriearter, der har fået hele deres genom sekventeret, fordelt på 7 phyla (se appendiks). De undersøgte arter repræsenterer et bredt udsnit af flagelbærende bakterier, fra fjernt beslægtede phyla, med mange forskellige fysiologier: Aquifex aeolicus er en termofil bakterie, der lever i vand tæt på kogepunktet, og som menes at være et af de ældste bakterie-phyla. Spirokæter har som de eneste bakterier hele flagellen inden for den ydre cellemembran, hvilket stiller unikke funktionelle krav. Borrelia burgdorferi har et lille genom på blot 851 gener, resultatet af kraftig reduktiv evolution. Der er med andre ord tale om arter, der er adskilt af milliarder af års evolution i vidt forskellige miljøer.
Ved hjælp af The Universal Protein Resource (UniProt) har jeg for hver af disse arter identificeret, hvilke flagelrelaterede gener den pågældende art har.
Et flagelrelateret gen, der findes i alle eller næsten alle flagelbærende arter, må nødvendigvis have været til stede i disse arters sidste fælles stamform, og må således have været del af ur-flagellen.
For at identificere ur-flagellens irreducible kerne har jeg for hvert gen undersøgt, om “knock-out” eksperimenter har påvist, at det pågældende gen kan fjernes uden tab af bakteriens evne til at bevæge sig. Hvis et gen kan fjernes, uden at bakterien mister sin evne til at bevæge sig, er dette gen ikke en del af den irreducible kerne.
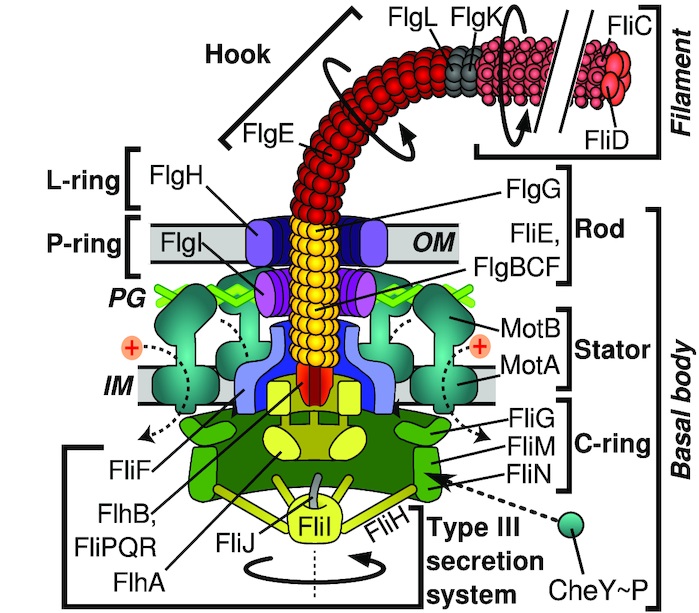
Jeg har således identificeret 25 flagelrelaterede gener, der sandsynligvis har været til stede i den sidste fælles stamform til alle flagelbærende bakterier, og dermed kan siges at udgøre “ur-flagellen”. Af disse gener klassificeres de 19 som værende del af flagellens irreducible kerne (tabel 1).
| Funktionsenhed | Gen | Del af kerne? |
| Svingtråd | fliC fliD | Ja Ja |
| Svingtråd/krog-samling | flgK flgL | Ja Ja |
| Krog | flgD flgE | Ja Ja |
| L-ring | flgH | Nej1 |
| P-ring | flgI | Nej1 |
| Aksel | flgB flgC flgF flgG fliE | Ja Ja Nej2 Ja Ja |
| Motor/Stator | motA motB | Ja Ja |
| MS-ring | fliF | Ja |
| C-ring/Rotor | fliG fliM fliN | Ja Nej3 Nej3 |
| Sekretionssystem | flhA flhB fliI fliP fliQ fliR | Ja Ja Nej3 Ja Ja Ja |
Kommentarer om udvalgte gener
- flgHI
Generne for FlgH og FlgI (der udgør henholdsvis L- og P-ringen, også kaldet bøsningerne) er til stede i alle undersøgte arter på nær repræsentanter for de gram-positive og for Spirochaetes. I gram-negative arter bruges bøsningerne til at forankre flagellen i den ydre cellemembran og peptidoglykanlaget. Hos gram-positive arter er disse lag fraværende, og der er dermed intet behov for bøsningerne. Hos de skrueformede Spirochaetes befinder hele flagellen sig inden for den ydre cellemembran. Flagellen går således ikke på tværs af den ydre cellemembran, hvorfor disse arter heller ikke har behov for bøsninger.
Fylogenetiske analyser peger på, at gram-negative bakterier, med deres to cellemembraner, er den ældste bakterietype, hvorfra gram-positive bakterier er opstået flere gange uafhængigt af hinanden gennem tab af den ydre cellemembran (Antunes et al. 2016, Taib et al. 2020).
Det er derfor sandsynligt, at bøsningerne har været til stede i ur-flagellen. Men da de ikke er til stede i alle de undersøgte arter, er de ikke klassificeret som del af flagellens irreducible kerne.
- flgF
Et enkelt gen (flgF), hvis genprodukt findes i akslen, mangler i tre af de undersøgte arter. Det er overraskende, at der ikke er større variation i akslens sammensætning, da den ‘bare’ er en stang, der overfører kraften fra motoren til svingtråden. At fire ud af fem af generne i akslen kan findes i alle arter, tyder på, at akslen er underlagt flere funktionelle krav, end man umiddelbart skulle tro.
Genet flgF har sandsynligvis været til stede i ur-flagellen (Liu & Ochman 2007), men da det ikke findes i alle de undersøgte arter, er det ikke klassificeret som del af flagellens irreducible kerne.
- fliI
Genet fliI (der koder for en ATPase i flagellens eksportsystem) er til stede i alle undersøgte arter og må sandsynligvis have været del af ur-flagellen. Ikke desto mindre har laboratorieforsøg vist, at bakterier udviser svag bevægelse, selv hvis de mangler fliI (Paul et al. 2008; Minamino & Namba 2008). Genet er derfor ikke klassificeret som del af flagellens irreducible kerne.
- fliMN
Generne fliM og fliN (der koder for to af de tre genprodukter, som C-ringen består af) er til stede i alle undersøgte arter og må sandsynligvis have været del af ur-flagellen. Ikke desto mindre har laboratorieforsøg vist, at bakterier uden de to gener udviser bevægelser, hvis genernes fravær kompenseres af en opregulering af de transkriptionsregulerende gener flhC og flhD (Erhardt & Hughes 2010).
Man kan forestille sig, at genprodukterne for fliM og fliN fungerer som en tragt, der leder materialerne fra sekretionssystemet op gennem flagellen. Når tragten mangler, kan flagellen ikke blive konstrueret, medmindre man “skruer op for hanen” ved at opregulere produktionen af de materialer, der skal gennem tragten.
Dette er en højst ineffektiv måde at konstruere flagellen, idet størstedelen af materialer går tabt, og det er tvivlsomt, om der nogensinde har eksisteret flageller uden denne tragtfunktion. Men strengt taget er den ikke nødvendig for flagellens evne til at udføre sin primære funktion, og derfor klassificeres fliM og fliN ikke som del af flagellens irreducible kerne.
Irreducibel kompleksitet som udfordring for evolutionære forklaringer
I det ovenstående har jeg identificeret de 19 gener, der udgør den bakterielle flagels irreducible kerne. Dette gør os i stand til at kvantificere den udfordring, den bakterielle flagel udgør for ethvert evolutionært forsøg på at forklare dens oprindelse.
Observationen, at flagellen er irreducibelt kompleks, udelukker i sig selv en række evolutionære forklaringer.
Vi kan således afvise, at flagellen er et produkt af af en gradvis, direkte evolutionsproces (Thornhill & Ussery 2000; se også Krauze 2020c).
Et eksempel på en gradvis, direkte evolutionsproces er Charles Darwins forsøg på at forklare øjets udvikling i Arternes Oprindelse (1859, s. 217-9). Darwin foreslog, at øjet oprindeligt blot bestod af en lysfølsom plet, der kun kunne skelne lys fra skygge. Pletten blev gradvist til en lysfølsom fordybning, så organismen nu kunne skelne, fra hvilken retning lyset kom. Den lysfølsomme fordybning blev gradvist fyldt af et geleagtigt materiale, der fungerede som en linse. Processen var ikke blot gradvis, men også direkte: Gennem hele processen havde systemet den samme funktion, som det blev bedre til at udføre.
Eller tag det klassiske scenarie med giraffen, der over generationer gradvist kunne nå højere og højere placerede blade, fordi dens hals gradvist blev længere og længere: En oprindelig struktur blev gradvist bedre til at udføre sin oprindelige funktion.
Vi kan afvise alle sådanne forklaringer, når det gælder den bakterielle flagel. Hvordan flagellen end er opstået, er den ikke opstået ved, at en proto-flagel bestående af en enkelt del gradvist øgede bakteriens evne til at bevæge sig, efterhånden som flere dele blev tilføjet. En flagel, der mangler en af delene, der udgør dens irreducible kerne, gør ikke bakterien i stand til at bevæge sig overhovedet.
Vi kan også afvise den forklaring, at flagellen opstod ved en springvis evolutionsproces, hvor en enkelt heldig mutation ved et rent lykketræf samlede hele systemet. Et sådant scenarie kan muligvis forklare et meget simpelt irreducibelt komplekst system, bestående af 2-3 dele, men næppe et system bestående af 19 nødvendige dele.
Hvad med en gradvis, indirekte evolutionsproces, hvor et system oprindeligt har én funktion, men bliver tilpasset til at udføre en anden funktion? Denne forklaring kan ikke principielt afvises. Men hvad er sandsynligheden for, at et system, der igennem generationerne er blevet tilpasset én funktion, tilfældigvis har en udformning, der gør det velegnet til i stedet at udføre en helt anden funktion? Som evolutionsbiologen Allan Orr, der ellers er stærkt kritisk over for Behes argument, bemærker:
“… vi kan muligvis tænke os, at nogle af delene i et irreducibelt komplekst system udviklede sig skridt for skridt med et andet formål for øje og så i stor stil blev rekrutteret til at udføre en ny funktion. Men dette er også usandsynligt. Man kan lige så vel håbe at halvdelen af ens bils gearkasse pludselig vil hjælpe til med airbaggen. Sådanne ting kan muligvis finde sted meget, meget sjældent, men de udgør bestemt ikke en generel løsning til irreducibel kompleksitet.” (Orr 1996)
Identifikationen af delene i flagellens irreducible kerne gør os i stand til bedre at evaluere sådanne indirekte scenarier ved at rette vores fokus mod de dele, hvis oprindelse scenariet skal kunne forklare.
I en kommende artikel vil jeg kaste et kritisk blik på det mest populære indirekte scenarie – at den bakterielle flagel udviklede sig fra det såkaldte type III-sekretionssystem, også kendt som injektisomet.
Designtilhængeren, der ønsker at afdække, om den bakterielle flagel er et produkt af intelligent design, står i en lignende situation som en evolutionsbiolog, der skal afdække, om et givent biologisk træk skyldes naturlig selektion eller genetisk drift (Sober, 2008). Her kan man ikke konkludere, at blot fordi den ene forklaring er en mulighed, er der ingen grund til at overveje den anden. På samme måde behøver designtilhængere ikke at udelukke enhver tænkelig og utænkelig evolutionær forklaring for at overveje den mulighed, at flagellen er designet.
Irreducibel kompleksitet er et redskab, der lader os fokusere på de systemer, hvor en evolutionær forklaring møder de største udfordringer, og som er kandidater til at være designede. Med identifikationen af den bakterielle flagels irreducible kerne er en designbaseret undersøgelse af systemet nået et skridt videre.
Appendiks: Oversigt over arter brugt i identifikationen af ur-flagellen og dennes irreducible kerne
| Gruppe | Art | Mangler? |
| Aquificae | Aquifex aeolicus | |
| Bacteriodetes | Salinibacter ruber | flgF |
| Firmicutes | Bacillus subtilis Clostridium tetani Listeria innocua | flgH, flgI flgH, flgI, flgF flgH, flgI, flgF |
| Planctomycetes | Rhodopirellula baltica | |
| Spirochaetes | Treponema denticola Borrelia burgdorferi | flgH, flgI flgH |
| Thermotogae | Thermotoga maritima | |
| α-proteobacteria | Caulobacter vibrioides | |
| β-proteobacteria | Chromobacterium violaceum | |
| γ-proteobacteria | Escherichia coli | |
| δ-proteobacteria | Desulfovibrio vulgaris | |
| ε-proteobacteria | Campylobacter jejuni |
Referencer
- Antunes L.C.S. et al., 2016, “Phylogenomic analysis supports the ancestral presence of LPS-outer membranes in the Firmicutes”, eLife 5:e14589. https://doi.org/10.7554/eLife.14589.001
- Beeby M., Ferreira J.L., Tripp P., Albers S.V., Mitchell D.R., 2020, “Propulsive Nanomachines: The Convergent Evolution of Archaella, Flagella and Cilia”, FEMS Microbiology Reviews 44(3):253-304. https://doi.org/10.1093/femsre/fuaa006. PMID: 32149348
- Behe M.J., 1996, Darwin’s Black Box: The Biochemical Challenge to Evolution, Simon & Schuster.
- Bock W.J., 2000, “Explanatory History of the Origin of Feathers,” Integrative and Comparative Biology 40(4):478-85. https://doi.org/10.1668/0003-1569(2000)040[0478:EHOTOO]2.0.CO;2
- Chen S. et al., 2011, “Structural diversity of bacterial flagellar motors”, The EMBO Journal 30(14):2972-81. https://doi.org/10.1038/emboj.2011.186
- Darwin C., 1859, The Origin of Species, republication of the first edition, Gramercy Books.
- Deckert G. et al., 1998, “The complete genome of the hyperthermophilic bacterium Aquifex aeolicus“, Nature 392(6674):353-8. https://doi.org/10.1038/32831
- Dembski W.A., 2002, No Free Lunch: Why Specified Complexity Cannot Be Purchased without Intelligence, Rowman & Littlefield.
- DeRosier D.J., 1998, “The Turn of the Screw: The Bacterial Flagellar Motor”, Cell 93(1):17-20. https://doi.org/10.1016/s0092-8674(00)81141-1
- Diepold A. & Armitage J.P., 2015, “Type III Secretion Systems: The Bacterial Flagellum and the Injectisome”, Philosophical Transactions of the Royal Society B 370(1979):20150020. http://doi.org/10.1098/rstb.2015.0020
- Erhardt M., Hughes K.T., 2010 “C-ring Requirement in Flagellar Type III Secretion is Bypassed by FlhDC Upregulation”, Molecular Microbiology 75(2):376-93. https://doi.org/10.1111/j.1365-2958.2009.06973.x
- Jones D., 2008, “Engines of Evolution”, New Scientist 197(2643):40-3. https://doi.org/10.1016/S0262-4079(08)60422-4
- Krauze M., 2020a, “Fælles design og hexameriske helicaser”, Intelligent Design DK. https://intelligentdesign.dk/2020/04/04/faelles-design-og-hexameriske-helicaser/
- Krauze M., 2020b, “Design og det første livs non-hierarkiske mønster”. https://intelligentdesign.dk/2020/08/29/design-og-livets-non-hierarkiske-monster/
- Krauze M., 2020c, “Hvad irreducibel kompleksitet kan – og ikke kan – bruges til”, Intelligent Design DK. https://intelligentdesign.dk/2020/12/12/hvad-irreducibel-kompleksitet-kan-og-ikke-kan-bruges-til/
- Liu R., Ochman H., 2007, “Stepwise formation of the bacterial flagellar system”, PNAS 104(17):7116-21. https://doi.org/10.1073/pnas.0700266104
- Macnab R.M., 2003, “How Bacteria Assemble Flagella”, Annual Review of Microbiology 57(1):77-100. https://doi.org/10.1146/annurev.micro.57.030502.090832
- Minamino T., Namba K., 2008, “Distinct roles of the FliI ATPase and proton motive force in bacterial flagellar protein export”, Nature 451(7177):485-8. https://doi.org/10.1038/nature06449
- Moens S., Vanderleyden J., 1996, “Functions of Bacterial Flagella”, Critical Reviews in Microbiology 22(2):67-100. https://doi.org/10.3109/10408419609106456
- Nakamura S., Minamino T., 2019, “Flagella-Driven Motility of Bacteria”, Biomolecules 9(7):279. https://doi.org/10.3390/biom9070279
- Orr H.A., 1996, “Darwin v. Intelligent Design (Again): The latest attack on evolution is cleverly argued, biologically informed – and wrong”, Boston Review 22 (6). https://bostonreview.net/archives/BR21.6/orr.html
- Pallen M.J., Penn C.W., Chaudhuri R.R., 2005, “Bacterial Flagellar Diversity in the Post-Genomic Era, Trends in Microbiology 13(4):143-9. https://doi.org/10.1016/j.tim.2005.02.008. PMID: 15817382
- Paul K., Erhardt M., Hirano T. et al., 2008, “Energy Source of Flagellar Type III Secretion”, Nature 451:489-92. https://doi.org/10.1038/nature06497
- Perlman M., 2009, “Traits Have Evolved to Function the Way They Do Because of a Past Advantage”, i Ayala F.J. & Arp R. (red.), 2009, Contemporary Debates in Philosophy of Biology, Wiley-Blackwell.
- Ruse M., 2003, Darwin and Design: Does Evolution Have a Purpose?, Harvard University Press.
- Santiveri M. et al., 2020, “Structure and Function of Stator Units of the Bacterial Flagellar Motor”, Cell 183(1):244-57. https://doi.org/10.1016/j.cell.2020.08.016
- Snyder L.A.S., Loman N.J., Fütterer K., Pallen M.J., 2009, “Bacterial flagellar diversity and evolution: seek simplicity and distrust it?”, Trends in Microbiology 17(1):1-5. https://doi.org/10.1016/j.tim.2008.10.002
- Sober E., 2008, Evidence and Evolution: The Logic Behind the Science, Cambridge University Press.
- Sober E. & Steel M., 2002, “Testing the Hypothesis of Common Ancestry”, Journal of Theoretical Biology 218(4):395-408. https://doi.org/10.1006/jtbi.2002.3086
- Taib N. et al., 2020, “Genome-wide analysis of the Firmicutes illuminates the diderm/monoderm transition”, Nature Ecology and Evolution 4(12):1661-72. https://doi.org/10.1038/s41559-020-01299-7
- Thain M., Hickman M., 2000, The Penguin Dictionary of Biology, tenth edition, Penguin Books.
- Thomas J., 1999, “Flagellar Variations, Minimal Complexity, and Evolutionary Noise”, Intelligent Design Theory. http://web.archive.org/web/20010722100738/http://www.idtheory.net/enoise.html. Oprindelig postet i talk.origins: https://groups.google.com/g/talk.origins/c/6ixm8YDqYKQ/m/GauBxYzxn28J
- Thornhill R.H. & Ussery D.W., 2000, “A Classification of Possible Routes of Darwinian Evolution”, Journal of Theoretical Biology 203(2):111-6. https://doi.org/10.1006/jtbi.2000.1070
Kategorier
